Ağrı Duyusuna Evrimsel Bakış
An Evolutionary Perspective to the Pain Sense
Hazal Artuvan KorkmazUzm. Dr., Fizyoloji Anabilim Dalı, Tıp Fakültesi, Ankara Üniversitesi, Ankara
[email protected]
Özet
Ağrı, insanlar için “gerçek veya potansiyel doku hasarıyla ilişkili hoş olmayan duyusal ve duygusal bir deneyim” ve diğer canılar için “caydırıcı bir duyu” olarak tanımlanır. Ağrı kelimesi birçok dilde işkence çekmek, bir şeyin cezasını çekmek gibi olumsuz duygularla ilişkilendirilmiştir. Ağrı duyusu mekanik, termal, kimyasal veya her üç uyarı tipine özelleşmiş nosiseptörler (ağrı almaçları) tarafından kodlanır. Filogenetik olarak farklı organizasyonlar gösteren duyu yolları ile beyne iletilir. Duyu korteksine iletilen informasyon işlenir; bilinçli duyu olarak canlı tarafından algılanır. Limbik sisteme iletilen informasyon ise ağrıya verilen duygusal ve otonom yanıtta rol alır. Ağrı duyusunun hissedildiği şiddet veya süre, her zaman uyaranın şiddeti ve süresi ile korelasyon göstermez. Ağrı duyusu özneldir; ölçümünde anketler, davranışsal gözlemler ve fizyolojik parametreler değerlendirilir. İnsan dışı hayvanlarda kendini ifade etmeden yoksunluk ve sinir sistemlerinin gelişkinliğindeki farklılıklar, bu canlıların bilinçli bir ağrı duyusuna sahip olup olmadıkları sorusunu sıklıkla sordurmuştur. Bu çalışma, omurgalı ve omurgasızlardaki çalışmaları özetleyerek ağrı duyusunun evrimsel avantajlarını ve dezavantajlarını tartışmayı amaçlamaktadır. Memelilerde ve omurgasızlarda nosisepsiyonla ilgili işlevsel paralellikler görülmüştür. Bu durum nosiseptif yanıtların yarım milyar yıldan fazla bir süredir korunduğunu ve/veya bu modelin yakınsak evrimin bir ürünü olduğunu düşündürmektedir. Nosisepsiyon varlığı her zaman bilinçli duyuyu göstermez. Çalışmalarda, genel olarak omurgalıların tümünün ağrı duyusunun farkında olduğu, omurgasızların ise bilinçli ağrıdan yoksun olduğu kabul edilmiştir. Bununla beraber, omurgasızlar sınıfındaki bazı canlı türlerinin ağrıya stres yanıtları ve sakınma davranışı geliştirdikleri bildirilmiştir. Ağrı duyusunun, klinik ve yaşamsal önemine ek olarak etik önemi de bulunmaktadır. Bu nedenle ağrı duyusuna geniş bir perspektiften ve evrimsel açıdan bakmak değerlidir.
Anahtar kelimeler: Ağrı Duyusu, Ağrının Evrimi, Nosisepsiyon.
Abstract
Pain is defined as an “unpleasant feeling and emotional experience associated with actual or potential tissue damage” for humans and “an aversive sensation” for other animals. In many languages, the word of pain has been associated with bad feelings such as suffering or punished for something. Pain sensation is encoded by nociceptors specialized for mechanical, thermal, chemical, or all three types of stimuli. It is transmitted to the brain by sensory pathways that have phylogenetically different organizations. The information transmitted to the sensory cortex is perceived consciously. The information transmitted to the areas of the limbic system has a role in the emotional and autonomic response to painful stimuli. The intensity or duration of the pain sensation does not always correlate with the intensity and duration of the stimulus. Pain sensation is highly subjective, and the experiments that aim to measure pain sensation use questionnaires, behavioral observations, and physiological parameters. The lack of self-expression in non-human animals and the differences in the development level of their nervous systems often raise the question of whether these creatures have a conscious sense of pain. This study discusses the evolutionary advantages and disadvantages of pain sensation by summarizing studies in vertebrates and invertebrates. Functional similarities in nociception have been seen in mammals and invertebrates. This indicates that nociceptive responses have been conserved for more than half a billion years and/or that this pattern is a product of convergent evolution. The presence of nociception does not always indicate the conscious pain sense. In the studies, it has been accepted that all vertebrates are aware of the sensation of pain, while invertebrates may not be aware of pain consciously. However, some species in the invertebrate class have also been reported to develop stress responses to pain and avoidance behavior. The sense of pain has clinical and vital importance, and it also has ethical importance. Therefore, it is essential to look at the sensation of pain from a broad and evolutionary perspective.
Key words: Pain Sense, The Evolution of Pain, Nociception.
Ağrının Tanımı ve Ağrı Fizyolojisi
Klinik tanıma göre ağrı “gerçek veya potansiyel doku hasarıyla ilişkili hoş olmayan duyusal ve duygusal bir deneyim”dir (Uluslararası Ağrı Çalışmaları Derneği) (The International Association for the Study of Pain (IASP) , 2021). Biyolojik tanıma göre ise “gerçek veya potansiyel doku hasarıyla ilişkili caydırıcı bir duyu” olarak ifade edilir (Broom, 2001). Klinik tanımda geçen duygu kavramının farkındalık gerektirmesi ve bunun her canlıya uyarlanmasının zor olmasından dolayı tanımda olmaması gerektiği düşünülmektedir. Biyologların tanımında ağrının öğreticiliği sıkça vurgulanır ve canlının ağrı duyduğunu varsaymak için öğrenilmiş kaçınmayla sonuçlanması ve türe özgü bir davranış geliştirmesi gerektiği savunulur, buna refleksler dâhil edilmez.
Tanımı daha da açabilmek için ağrının sözcük kökenine bakmak da faydalı olacaktır. İngilizce karşılığı olan painkelimesinin Anglo-Fransız peine (pain, suffering – işkence), Latince poena (penalty, punishment - ceza, cezalandırma) ve Yunanca poine (payment, penalty, recompense - ödeme, ceza, tazminat) sözcüklerinden köken aldığı düşünülür (Wictionary The Free Dictionary, 2021). Türkçede ise Uygur ve Orhun yazılarında agrıg (hastalanmak, acı çekmek) sözcüğü şeklinde kullanılmıştır (Nişanyan, 2021). Ağız sözcüğünün türevi olduğu ve özgün anlamın “haykırmak, avaz etmek”ten geliyor olabileceği belirtilmektedir. Görüldüğü gibi, insanlar tarih boyunca ağrı duyusunu işkence çekme, ceza ödeme şeklinde olumsuz duygu yükleri ile ifade etmiştir.
Ağrı duyusuyla beraber sıklıkla karşımıza çıkan bir diğer terim olan nosere ise fizyolojik anlamda ağrı duyusundan farklı değerlendirilmelidir. Latince kökeni nocere (to hurt – canını yakmak, acıtmak) olan bu sözcük ağrı duyusu reseptörlerine de ad vermiştir (nosiseptör).
Reseptörler (almaçlar), belli enerji formlarına maksimum yanıt veren, özelleşmiş hücrelerdir. Örneğin, mekanik enerjiye duyarlı mekanoreseptörler, kimyasal enerjiye duyarlı kemoreseptörler ve termal enerjiye duyarlı termoreseptörler tanımlanmıştır. Ağrı duyusu için bu üç enerji tipi de önemlidir. Hem mekanik hem sıcak (>45 derece) hem soğuk (<5 derece) hem de bazı kimyasallar (5-hidroksitriptamin, histamin, prostaglandinler, bradikinin, asidik pH, sitokinler ve tripsin gibi bazı enzimler) için özelleşmiş nosiseptörler bulunur (Mertens ve ark., 2015). Bunların yanı sıra diğer duyu modalitelerinden farklı olarak, her üç uyaran tipine de yanıt veren polimodal nosiseptörler vardır. Duyu sistemlerinde uyaranın modalitesi, konumu, şiddeti ve süresi reseptör düzeyinden itibaren kodlanmaya başlar. Ağrı duyusu nöral işlemleme sırasındaki bir takım merkezi ve çevresel düzenlemelerin sonucunda, daha uzun veya kısa süreli, daha az (analjezi) veya çok (hiperaljezi) şiddetli algılanabilmektedir. Ayrıca getirici (aferent) duyu nöronlarının anatomisi nedeniyle, kaynaklandığı bölge dışında bir yerde oluşabilmekte (ör. yansıyan ağrı), hatta ağrı verici uyaran yokluğunda bile ağrı hissedilebilmektedir (ör. allodini, fantom ağrı). Örneğin ağrı duyusunun aferent nöronlarında görülen akson refleksi neticesinde zarar verici uyaran ortadan kalksa bile, çevre dokudaki mast hücrelerin aktivasyonu ve artan histamin nedeniyle polimodal reseptörler tekrardan uyarılır; ağrı daha yaygın, uzun süreli ve şiddetli hissedilmeye devam eder (Mtui, 2016). Ağrı duyusunun kodlanmasında etkisi gösterilen birçok nöromodülatör bulunur (P maddesi, CGRP, histamin, endojen opioitler). Endojen opioitler, ağrı duyusunu baskılayıcı nitelik taşır ve ağrının yukarıdan aşağıya nöral düzenlemesinde rol alır. Merkezi modülasyonda limbik sistem de önemlidir, önceki deneyimler veya motivasyonun duyulan ağrı şiddetini etkilediği bilinmektedir (Ör. bir çatışma sırasında yaralanan askerin bunu ancak çatışma bittiğinde fark etmesi) (Barrett ve ark., 2019).
Aferent ağrı lifleri tüm aferent duyu aksonları arasındaki en ince ve miyelinsiz lifler olan A delta ve C tipi liflerdir. Bu sebeple ağrı (ve bazı termal duyular) en yavaş iletilen duyu modalitesidir (Smith ve Gary, 2009). A delta tipi lifler hızlı, keskin, daha iyi lokalize edilebilen ağrıdan sorumludur, yavaş adaptasyon (uyum) gösterir. Ağrıya verilen geri çekme refleksi ve haykırma yanıtına neden olur. C tipi lifler, tüm aferent lif tipleri arasında en fazla bulunan lif tipidir ve gecikmiş, yavaş ağrıdan sorumludur, adaptasyon göstermezler (Smith ve Gary, 2009; Stevens, 2009). C tipi liflerle merkeze iletilen yavaş ağrıya sıklıkla otonom belirtiler eşlik eder. En ince lif tipi olmasının metabolik bir avantaj sağladığı ve aynı zamanda kolay uyarılabilmeye olanak sağladığı düşünülebilir.
Ağrıyla ilişkili duyu yolları evrimsel olarak farklı zamanlarda oluşan filogenetik yapılanma gösterir (Smith ve Gary, 2009). Şekil 1’de gösterildiği gibi, nosiseptif sinyallerin bir kısmı evrimsel olarak daha yeni olan neospinotalamik yol üzerinden somatik duyu korteksine iletilir. Neospinotalamik yol somatotopik yapılanma gösterir. Bu yolda iletilen ağrının kaynaklandığı vücut bölgesi daha iyi lokalize edilir. Nosiseptif sinyallerin bir kısmı ise daha eski olan paleospinotalamik yol üzerinden anteriyor singulat girus ve insula gibi limbik sistem alanlarına iletilir. Bu yol, ağrıya verilen duygusal ve otonom yanıta katılır. Ayrıca ağrı informasyonu paleospinotalamik yol üzerinden mezensefalon periakuaduktal gri madde ve uyku-uyanıklığı düzenleyen retiküler formasyon çekirdeklerine iletilir. Bu yol endojen analjeziden de sorumludur (Barrett ve ark., 2019). Endojen analjezi, bir takım vücut içi mekanizmalarla ağrının azaltılması anlamına gelir.
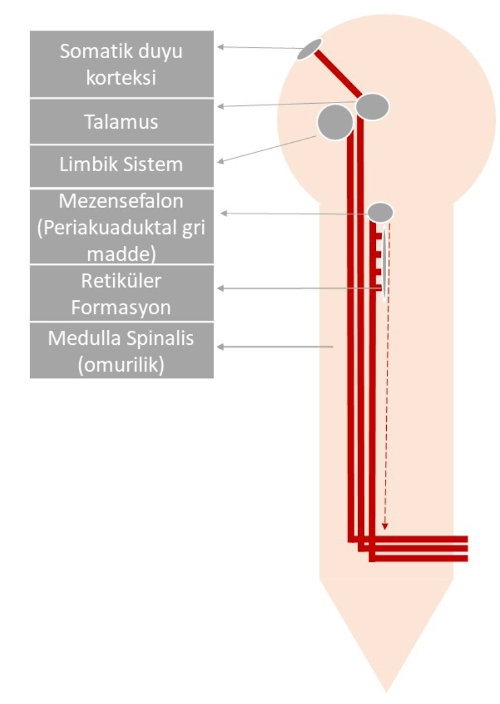
Şekil 1: Ağrı duyusu yollarının anatomisi. Nosiseptörlerden gelen bilgi önce omuriliğe daha sonra projeksiyon lifleri (kalın çizgiler) ile beyne ulaşır. Bu projeksiyon nöronlarının bir alt kümesi, bilgiyi talamus yoluyla somatik duyu korteksine ileterek ağrılı uyaranın yeri ve ağrının şiddeti hakkında bilgi sağlar. Diğer projeksiyon nöronları, beyin sapındaki ve limbik sistemdeki bağlantılar yoluyla ağrı deneyiminin duygusal bileşenine katkıda bulunur. Bu bilgi aynı zamanda periakuaduktal gri madde nöronlarına da erişerek omurilikten çıkan yolları düzenleyen inen yolları (kesikli çizgi) devreye sokar. Periakuaduktal gri maddeden inen lifler, merkeze çıkan yollar üzerinde baskılayıcı etki yaparak duyulacak ağrının şiddetini azaltır.
Canlı türlerinde ağrı duyusu incelenirken dikkat edilmesi gereken noktalardan biri, bazı canlıların sinir sisteminin duygusal yanıt oluşturacak ve farkındalık atfedilecek düzeyde gelişmemiş olmasıdır. Canlıların ağrı duyup duymadığı konusunda bazı kısıtlılıklarımızın olduğunu kabul ederek ilerlememiz gerekir. Örneğin, insanlarda dahi ağrı duyusu, yukarıda bahsedilen fizyolojik mekanizmalar nedeniyle oldukça özneldir ve uyaran şiddetiyle her zaman korele değildir (Petersen-Felix ve Arendt-Nielsen, 2002). Bu bağlamda ölçülebilirliği de tartışmalıdır. İnsanlarda ağrı ölçekleri ile değerlendirme yapılabilirken diğer hayvan türlerinde dışarıdan gözlemlenebilen davranışsal veya fizyolojik değişikliklerden yararlanılır (mimik, sakınma davranışı, nabız, kortizol, elektrofizyoloji gibi) (Sneddon ve ark., 2014). Bu teknikler, omurgasızlardaki sinir sistemi farklılıkları nedeniyle kısıtlıdır.
Türler Arasında Nosiseptörlerdeki Benzerlikler
Bakteriler sinir sistemi bulunmayan ilkel tek hücreliler olmalarına rağmen zarar verici uyaranı saptayabilirler. Örneğin, Escherichia coli.de bulunan bir tür mekanik reseptörün lizisten korunma mekanizması olarak geliştiği öne sürülmüştür. Aşağıda gelişkinlik seviyesi artacak şekilde birkaç canlı türü nosiseptörler açısından irdelenmiştir: Deniz şakayığının (sea anemonae) mekanik ve elektrik şokuna karşı kapanma refleksi ile yanıt verdiği, ancak termal duyarlılığının bulunmadığı gösterilmiştir (Smith ve Gary, 2009). Bu durumun, birlikte yaşadığı yengeçlerden sakınma davranışı olabileceği düşünülmüştür. Kapanma refleksi nosiseptif duyunun varlığını gösterse de bu canlı merkezi sinir sisteminden yoksun olduğundan ağrı duyusu anlamına gelmediği söylenebilir. Halkalı solucan (annelida) ganglionlarında 3 tip sinir hücresi tanımlanmıştır: dokunma, basınç ve nosiseptör. Her birinden alınan elektrofizyolojik kaydın memelilerdekine benzediği bildirilmiştir (Sneddon ve ark., 2014). Genel olarak omurgasızlardaki nosiseptörlerden şiddetli basınç, delinme, sıkışma veya yanma sırasında alınan hücre içi kayıtlar, diğer duyu nöronlarına göre yavaş ileten, küçük çaplı, tekrarlanan uyarım ile yanıt şiddeti fazla değişmeyen, yani yavaş adapte olan aferentlerin memeli nosiseptörüne benzediğini göstermiştir. Aynı zamanda opioid reseptörlerinin, dolayısıyla analjezi mekanizmasının varlığı da bildirilmiştir. Bakteri ile enfekte edilen bir kelebek larvasında hastalık davranışına eşlik eden allodini/hipersensivite benzeri bir durum gözlenmiştir (Adamo ve McMillan, 2019). Bu bulgu, bir hayvanın enfeksiyon nedeniyle zayıfladığında yırtıcılara karşı uyanıklığı sürdürmek için en ufak uyaranla bile çekilme davranışı oluşturduğu şeklinde yorumlanmıştır.
Omurgalı hayvanların çoğunun benzer ağrı reseptörlerine ve ilişkili merkezi sinir yollarına sahip olduğu bilinmektedir. Balık nosiseptörlerinin elektrofizyolojik özellikleri, daha yüksek omurgalılarda bulunanlarla hemen hemen aynıdır (Stevens, 2009; Sneddon ve ark., 2003). Bu canlılar arasında P maddesi, serotonin, kalsitonin gen ilişkili peptid, nöropeptid Y, bombesin ve metenkefalin gibi ağrı mediyatörlerinin ortaklığı gösterilmiştir (Cameron, 1990; Gregory, 1999; Sneddon ve ark., 2014). Memelilerde bulunan asit nosisepsiyonunda rol alan iyon kanalı (acid sensing ion channels; ASICs) genleri deniz kestaneleri, denizyıldızı, meşe palamudu solucanları ile memeli dışındaki omurgalılarda da gösterilmiş; bu genler evrimsel süreçte korunarak günümüze kadar gelmiştir (Sneddon, 2019). Yine memelilerde tanımlanan termal ve mekanik nosisepsiyonda görevli geçici reseptör potansiyel kanalı (transient receptor potential channels; TRP) genleri omurgasızlardan sirke sineğinde gösterilmiştir. Opioidler, merkezi sinir sisteminden inen yollarla ağrının baskılanmasını sağlayan (endojen analjezi), balıklarda ve memelilerde benzer mekanizmalarla çalışan morfin benzeri maddelerdir. Sirke sineğinde opioid reseptör genleri gösterilememiş olsa da dışarıdan opioid uygulanması ile analjezi elde edilmekte, ancak mekanizması henüz bilinmemektedir (Sneddon, 2019).
Sneddon ve ark. (2014) çok hücreli canlılardan itibaren nosiseptörlerin varlığı ve özelliklerinin derlendiği bir çalışma yayınlamıştır. Buna göre, sölenterlerde (cnidaria) mekanik nosiseptörlerin varlığı gösterilmiştir. Böceklerde (artropod), yuvarlak (nematod) ve halkalı (annelida) solucanlarda buna ek olarak sıcak ve kimyasal duyarlı nosiseptörler ve miyelinsiz C tipi lifler bulunmaktadır. Yumuşakçalarda (mollusca) mekanik nosiseptörler ve miyelinsiz ağrı lifleri vardır.
Omurgalılar sınıfındaki taşemenlerde (petromizon), mekanik ve sıcak nosiseptörleri ile bağlantılı miyelinsiz lifler olduğunu, miyelinli aferent liflerin ise ilk kez kemikli balıklarda (teleost) karşımıza çıktığını görüyoruz (Sneddon ve ark., 2014; Sneddon ve ark., 2003). Amfibiler ve sürüngenler mekanik, sıcak ve kimyasal nosiseptörlere sahiptirler. Memelilere gelindiğinde ise bunlara ek olarak soğuk nosiseptörleri vardır (Sneddon ve ark., 2014). Kuşlar ve memelilerde termoregülasyonun önemi düşünüldüğünde, organizmanın vücut sıcaklığını değiştirebilecek soğuk ya da sıcak bölgelerden kaçınmasında termal nosiseptörlerinin evrimsel önemi olduğunu söyleyebiliriz. Soğuk sularda yaşayan balıklarda soğuk nosiseptörü bulunmaması, insanlarda kapsaisine yüksek duyarlılık varken, acı biberdeki tohumların saçılmasına aracılık eden kuşlarda bu duyarlılığın olmaması gibi adaptif modülasyonlar bulunmaktadır (Sneddon ve ark., 2014). Günümüzden yaklaşık 635 milyon yıl önce sölenterlerin, 541 milyon yıl önceyse omurgalıların evrimleştiği (Futuyma ve Kirkpatrick, 2017) göz önüne alındığında, memelilerde ve omurgasızlarda nosisepsiyonla ilgili işlevsel paralellikler bize nosiseptif yanıtların yarım milyar yıldan fazla bir süredir korunduğunu ve/veya bu modalitenin yakınsak evrimin bir ürünü olduğunu göstermektedir (Walters ve Williams, 2019).
Hangi Canlılar Ağrı Duyar?
Birçok omurgasızda nosisepsiyonun varlığı gösterilmiş olsa da bu canlıların ağrı duyup duymadığı sadece nosiseptörler ile kanıtlanamaz. Ağrı verici uyaran nosiseptörler tarafından saptandıktan sonra informasyon daha üst merkezlere iletilir ve işlenir. Yukarıda değinildiği gibi, bu merkezlerin bir kısmı bilinçli bir kısmı bilinçsiz duyularla ilişkilidir. İnsan beyninde evrimsel olarak farklı zamanlarda oluşmuş üç katmandan bahsedilir: İlkel beyin (beyin sapı ve beyincik), ara beyin (limbik sistem) ve üst beyin (beyin kabuğu veya korteks) (Kaya, 2015). Beyin sapı, istemsiz çalışan solunum, dolaşım, sindirim gibi temel yaşamsal işlevleri düzenler. Limbik sistem, içgüdüsel davranışları da içeren programlı davranışları kontrol eder (ör. koruma davranışı). Üst beyin, farklı enerji formlarının önce birincil duyu alanlarında tek tek tanımlandığı, sonra ikincil ve daha üst düzey alanlarda bu bilgilerin birleştirilip bütünleştirildiği, böylece nesnel gerçeğin ayrıntılı bir imajının oluşturulduğu beyin katmanıdır. Üst beynin daha ilkel beyin bölümleri ile bağlantıları vardır, böylece yukarıdan aşağı ve aşağıdan yukarı süreçlerle hem bilinçli hem otomatik davranışlar şekillenir. Örneğin, keskin bir nesneyle ilk defa karşılaşan bir bebek bunun zararlı olabileceğini bilemez, ancak ağrı duyusunu deneyimledikten sonra bu bellek sistemlerinde depolanır ve öğrenilir. İleriki zamanlarda karşılaşılan keskin bir nesneye temas etme davranışını şekillendirir. Sosyal davranışlarda da benzer öğrenme mekanizmaları kurulabilir. Böylece, duyuların bilinçli olarak algılandığı üst beyin, otomatik olmayan, düşünerek ve planlanarak gerçekleştirilen daha karmaşık davranışlardan ve entelektüel işlevlerden sorumlu tutulur. Diğer canlıların ağrıyı insanlardaki ağrı duyusu gibi deneyimleyip deneyimlemedikleri bu beyin devrelerindeki benzerliklerle ilişkilidir.
Hangi canlıların ağrı duyduğuna yanıt arayan çalışmalarda çoğunlukla ağrı verici uyarana karşı oluşan davranışlar incelenmiştir. Biyolojik tanıma döndüğümüzde ağrı duyduğu varsayılan canlının ağrı verici uyarana karşı türe özgü davranış geliştirmesi ve kaçınmayı öğrenmesi beklenir. Böceklerde, ağrı verici uyaran sonrası kısa-uzun süreli nöronal değişiklik olmadığı, öğrenme ve sakınma davranışı görülmediği, sadece geri çekme ve kaşıma gibi reflekslerin var olduğu bildirilmiştir (Eisemann ve ark., 1984). Mevcut bilgiler ışığında omurgasızların çoğunun ağrı hissetmediği kabul edilmiştir. Öte yandan, bu canlıların henüz tüm duyu mediyatörleri bilinmiyor, ağrılı uyarana karşı oluşturdukları ileti yöntemleri anlaşılamıyor olabilir. Gelecek çalışmalarda yeni yöntemlerle ağrı duydukları gösterilebilir. Dekapod artropodlarda (yengeç, kerevit) ağrılı elektrik uyaranı verildiğinde serotonin ve glukoz artışı ile karanlığı tercih etmede artış gözlenmiştir. Anksiyolitik verildiğindeyse memelilere benzer şekilde aydınlığı tercih etmede artış olmuştur (Sneddon ve ark., 2014; Patterson ve ark., 2007). Yengeçlere kabuktan ağrı oluşturacak şiddette elektrik verildiğinde kabuğu terk ettikleri gözlenmiş, ortama yeni bir kabuk konduğunda hızla bu yeni kabuğa girmeye çalıştıkları görülmüştür (Elwood ve Appel, 2009). Yeni bir kabuğa girme çabası kabuğun kendisini koruduğunu öğrenmesine bağlanabilir veya bu sadece genetik olarak kodlanmış bir davranış olabilir. Öte yandan, ağrı verici uyaranın öğreticiliği düşünüldüğünde, yeni kabuğa karşı sakınma davranışı sergilemediği de söylenebilir. Ağrının eğiticiliği ile ilgili bir örnek kalamarlar ile verilebilir. Ağrıyı deneyimleyen kalamarların sakınma davranışı gösterdiği ve yara almış kalamarların avcıdan çok daha uzak durduğu gözlenmiştir. Üstelik, ağrı kesici uygulandığında bu temkinli yaklaşma davranışı ortadan kalkmıştır (Crook, Dickson, Hanlon ve Walters, 2014). Yasalar önünde, bu hayvanların ağrısının farkında olduğu kabul edilmiştir ve hayvanlara yapılan zulümle ilgili mevzuata dahil edilirken, kafadanbacaklılar dışındaki omurgasızlar dahil edilmemiştir (The Senate Standing Committee on Legal and Constitutional Affairs, 2021).
Omurgalıların tümünün ağrı hissinin olduğu kabul edilmiştir. Balıklar, zararlı uyaranlara karşı güçlü otonom ve nöroendokrin stres yanıtları verir (Stephanie, 2008; Sneddon ve ark., 2003). Karşılaştırmalı nöroanatomik çalışmalar, balık beyninde diğer omurgalılarda gösterilmiş bazı kortikal sahaların yokluğu nedeniyle, balıkların ağrıya verilen duygusal yanıt veya korku gibi bir psikolojik kapasiteleri olmadığına işaret etmektedir (Rose, 2002). Memeliler, kuşlar ve sürüngenlerin tümü ağrıya duygusal tepkiler gösterir. Ağrılı uyarana karşı otonom yanıtlar oluşturur, türe özgü davranış geliştirir ve sakınmayı öğrenirler. Ağrı kesici ile bu yanıtlar azalır ve ağrı kesici kullanmayı tercih ederler (Sneddon ve ark., 2014). Bu da ağrıya bilinçli farkındalığı ve ağrının kurtulmak istenen bir duyu olduğunu düşündürür.
Sinir sistemleri gelişkin olan omurgalılarda bilinçli farkındalık daha yüksektir. Çoğunun yavru bakımı (koruma, temel yaşamsal becerileri öğretme) ve sosyal davranış geliştirdikleri (grup oluşturma, grup bireylerini tanıma, yardımlaşma) görülür. Primatlarda grup büyüklüğü ve sosyalleşme yüzdeleri ile bilincin evriminde rolü olduğu düşünülen neokorteks (yeni kabuk) büyüklüğü ilişkilidir (Dunbar, 1993). Neokorteks; bellek, karar verme, plan yapma ve sosyal davranışların ortaya çıkmasında rol alan ön beyin bölümüdür (Barrett ve ark., 2019). Omurgalıların bellek yetileri ve öğrenebilme kapasiteleri yüksektir, hayatta kalmak için ağrının omurgalılar için önemli bir öğrenme aracı olduğu düşünülmektedir. Tüm bu etmenler neticesinde omurgalılar daha uzun ömürlüdür (Broom, 2001).
Öte yandan, omurgasızlarda sinir sistemi o kadar gelişkin değildir. Sınırlı bilişsel kapasiteleri vardır. Yavru bakımı çoğunda yoktur hatta yavrularını yiyen türler bulunur. Çoğunun sosyal davranışı yoktur. Davranışları (avlanma, yırtıcılardan saklanma) genetik olarak belirlenir (Eisemann ve ark., 1984). Omurgasızların öğrenme kapasitesi sınırlıdır (ör. cama vuran sinek). Bu nedenle ağrının evrimi için evrimsel seçim baskısı daha az olabilir (Adamo, 2016). Caydırıcı bir uyarana karşı kaçış tepkisine sahip olmak, canlının ağrı hissederek bilinçli bir sakınma davranışı çıkardığını her zaman göstermez. Örneğin bakteriler de kaçabilir. Ağrı duymasa bile bakterilerin evrimsel uyumu çok yüksektir, o nedenle ağrı duyusu, her canlı türü için aynı evrimsel avantaj veya dezavantajı oluşturmaz.
Evrimsel avantaj sağlamayan bir işlev gereksiz yere enerji tükettiği için zamanla kaybolur. “Evrim Ekonomisi” gereği, işlevsiz uzuvların kaybolmasının sağladığı enerji tasarrufu ile daha işlevsel yapıların verimliliğinin arttığı ve bu sebeple evrimsel süreçte bazı organların körelmesinin avantaj sağladığı düşünülmektedir (Coşan, 2016). O halde bazı türler için ağrı duyusunun seçici bir avantajı olduğunu varsayabiliriz. Ağrı, hasara veya ölüme neden olabilecek eylemlerden kaçınmaya motive eder. Hasar verici uyaranın farkına varmak ve tedavi uygulamak söz konusu olduğunda ağrının bilince ulaşması yaşamsaldır. İnsanlardaki bir tür tirozin kinaz mutasyonu embriyonik dönemde aferent nöronların gelişmemesine ve ağrı duyarsızlığına yol açar. Ağrısızlık kulağa hoş gibi gelse de bu kişilerin yaşam beklentisi azalmıştır (Nagasako ve ark., 2003). Benzer şekilde, yara almış ancak analjezi uygulandığı için ağrı hissetmeyen kalamarların avcıdan uzak durma davranışı ortadan kalktığı için hayatta kalma oranları düşmüştür (Sneddon, 2019). Bunlar, ağrının ağrısızlıktan daha avantajlı olduğu durumlara örnektir. Büyük primatlar, köpekler ve domuzlar sosyal olarak yaşarlar, ebeveynler çocuklara yardım edebilir, hatta diğer grup üyeleri saldırıya uğrayan veya acı çeken kişilere yardım edebilir. Bir yaralanmadan kaynaklanan ağrı hissedildiğinde çığlık, bağırma gibi yüksek sesli tehlike sinyalleri iletişim amaçlı kullanılabilir. Ama bu canlılar ilginç bir şekilde doğum yaparken çok sessizdir (Broom, 2001; Finlay, 2015). Sessiz olmaları, doğumda ağrı hissetmedikleri anlamına gelmez. İnsanda hem doğum sancısı ile bağırmak hem de yenidoğan bebeğin ağlaması kendine yardım gelmesini sağlayarak hayatta kalmasını kolaylaştırmış olabilir. Ancak diğer memelilerde yenidoğanların yardım bekleme gibi bir durumu yoktur. Aksine yenidoğan bir memeli yavrusu ağlarsa yırtıcıları çağıracaktır. Bu durumda bazı memeli türlerinin yardım çağırmaktansa yırtıcıları çağırmaması daha avantajlı olacağından, evrimsel süreçte böyle bir davranış geliştirmiş olması mümkündür (Broom, 2001).
Ağrısız olmanın seçici avantajı da söz konusudur. Örneğin, çıplak köstebek-faresi deride ağrı hissi eksikliği sayesinde sert yer altı ortamında ve yüksek karbon dioksit seviyelerinde ağrı duymadan gelişebilir (Park ve ark., 2008). Güney çekirge fareleri, arizona akrebinin acı verici ve potansiyel olarak ölümcül sokmalarına karşı dirençlidir (Thompson, 2018). İnsan beyin dokusunda da nosiseptörler yoktur. Nöron gruplarının maksimum verimle, minimum hacimde çalışması avantajlı görünmektedir ve daha sıkı bir beyin yapısı lehine nosiseptörleri göz ardı etmek insanın evriminde bir avantaj sağlamış olabilir. Beynin evriminde kortikal katmanların ve girusların şekillenmesinin avantajından bahsedilirken, eklenen yeni nöron gruplarının kaplayacağı yer, harcayacağı enerji ve üreteceği ısının beynin evrimine kısıtlılık oluşturacağının altı çizilir (Coşan, 2016). Baş ağrısı, nöral dokunun kendisinden değil beyin zarları ve damarlarından duyu alan nosiseptörlerden kaynaklanmaktadır. İnsan beyninde nosiseptörlerin yokluğu metabolik ve kaplayacağı alan açısından avantaj sağlamış olsa da bugüne ulaşmayı başaran birçok genetik kökenli baş ağrısı hastalığı biliyoruz. Kronik ağrı genlerini günümüze getiren bir evrimsel avantaj söz konusu olabilir mi? Genetik geçişli ve oldukça yaygın görülen migren hastalığını ele alalım. Migrenle ilgili tanımlanan genlerin evrimsel bağlantıları henüz çözülemese de migrenin evrimsel bir avantajı olduğu lehine tartışmalar bulunmaktadır (Andreou ve Edvinsson, 2019). Migren, iç ve dış stres kaynaklarına bağlı bir yanıt olan geri çekilme ve uyuşukluk ile karakterizedir. Yoğun sese, ışığa, kokuya, fiziksel ve/veya zihinsel strese, uykusuzluğa, açlığa, alkol gibi tetikleyicilere tolerans azlığı görülür. Kişilerin ağrı atakları yaşamamak için çok düzenli ve tetikleyicilerden uzak bir yaşam sürmesi tavsiye edilir (Robblee ve Starling, 2019). Evrimsel olarak bakıldığında çevresel tehditleri derhal tespit etme ve bunlardan kaçınma, toksin alımından veya solunmasından kaçınma hayatta kalmayı artıracaktır. Bu özellikler, baş ağrısı nüksünün neden olduğu periyodik rahatsızlığa rağmen, yüksek adaptif değerleri nedeniyle doğal seçilim açısından avantaj sağlamış olabilir (Bonavita ve Simone, 2011).
Sonuç
Sonuç olarak ağrı duyusu hem klinik hem biyolojik bilimleri ilgilendiren bir modalitedir. Ağrının omurgalılar başta olmak üzere sosyal yaşayan birçok canlı için yaşamsal olduğu, ama bazen de ağrısızlığın yaşamsal ve evrimsel avantajı dikkat çekmektedir. İnsan biyolojisini temellendirmede evrimsel bağların kurulması, fizyolojik mekanizmaların, hastalıkların, ilaç yan etkilerinin veya tedaviye dirençlerin anlaşılmasında yararlı olabilir. Örneğin, klinikte sıkça karşılaşılan omurga kaynaklı ağrılar ve sinir sıkışmalarının, insanın iki ayak üzerinde durması ve alet kullanımı ile el-ayak eklemlerinin kullanış şekillerinin evrimsel uyum sağlanamadan hızla değişmesinin sonucu olduğu düşünülmektedir (Kaya, 2015). Doktorların tedavi etmeye uğraştıkları hastalıklar aslında bir tür evrimsel seçim baskısı ile mücadele olabilir veya migren örneğindeki gibi nesilden nesile aktarılarak türe fayda sağlayan bir genetik çeşitlilik söz konusu olabilir. Bugün tıpta nedeni bilinmeyen hastalık olarak etiketlenen birçok hastalık evrimsel uyum yasalarının sonuçları olabilir. Temel tıp literatüründe, eğitim ve pratiklerde evrimsel sürecin insan biyolojisini nasıl şekillendirdiği çoğunlukla tartışılmamaktadır. Bu konunun tartışılması için yeterli altyapı ve bilgi birikimi de yetersiz görülmektedir (Kaya, 2015). Hem fizyolojik hem patolojik durumlara evrimsel bakış açısı getirmek faydalıdır. Öte yandan, hangi canlılar ağrı duyar sorusunun etik önemi ve sosyal boyutu vardır. Gerek gıda sektöründe gerek deneysel araştırmalarda hayvanlara yapılan muamelelerde canlıların ağrı duyup duymaması, işlemin analjezi gerektirip gerektirmemesi gibi hususların göz ardı edilmemesi gerekir.
KAYNAKÇA
Adamo, S. A. ve McMillan, L. (2019). Listening to your gut: immune challenge to the gut sensitizes body wall nociception in the caterpillar Manduca sexta. . Phil. Trans. R. Soc. B, 374(20190278). http://dx.doi.org/10.1098/rstb.2019.0278.
Adamo, S. A. (2016). Do insects feel pain? A question at the intersection of animal behaviour, philosophy and robotics. Animal Behaviour, 118, 75-79.
Andreou, A. P. ve Edvinsson, L. (2019). Mechanisms of migraine as a chronic evolutive condition. The Journal of Headache and Pain 20:117. https://doi.org/10.1186/s10194-019-1066-0.
Barrett, K. E., Barman, S. M., Boitano, S. ve Reckelhoff, J. F. (2019). Ganong'un Tıbbi Fizyolojisi. (E. Ağar, ve İ. Alican) Güneş Tıp Kitabevleri.
Bonavita, V. ve Simone, R. D. (2011). Pain as an evolutionary necessity. Neurol Sci, 32(Suppl 1), 61-66.
Broom, D. (2001). The evolution of pain. Symposium on The science and Philosophy of Pain II, 70, s. 17-21. doi:10.1007/s10072-011-0539-y.
Cameron, A. P. (1990). Organization of the spinal cord in four species of elasmobranch fish: Cytoarchitecture and distribution of serotonin and selected neuropeptides. Journal of Comparative Neurology, 297, 201-218.
Coşan, E. (2016). Beyin ve bilinç evrimi. Osmangazi Tıp Dergisi, 38(1), 20-28.
Crook RJ, Dickson K, Hanlon RT ve Walters ET. (2014) Nociceptive sensitization reduces predation risk. Curr. Biol. 24,1121–1125. doi:10.1016/j.cub.2014.03.043.
Dunbar, R. (1993). Coevolution of neocortical size, group size and language in humans. Behavioral and Brain Sciences, 16(4), 681-694. doi:10.1017/S0140525X00032325.
Eisemann, H., Jorgensen, W. K., Merritt, D. J., Rice, M. J., Cribb, B. W., Webb, P. D. ve Zalucki, M. P. (1984). Do insects feel pain? — A biological view. Cellular and Molecular Life Sciences, 40(2), 164-167. doi:10.1007/BF01963580.
Elwood, R. W. ve Appel, M. (2009). Pain experience in hermit crabs? Animal Behaviour, 77, 1243–1246.
Finlay, B. (2015). The unique pain of being human. New Scientist (9 May 2015) sf.60. Erişim Tarihi: 27.05.2021 https://archive.org/details/New_Scientist_9_May_2015/page/n29/mode/2up.
Futuyma, D. J. ve Kirkpatrick, M. (2005). Evolution (4 b). Massachusetts: Sinauer Associates, Inc. s. 434.
Gregory, N. (1999). Do fish feel pain? ANZCCART News 12.
Kaya, T. (2015). Yaşam, Evrim ve Biz. İstanbul: Alfa Basım Yayım.
Mertens, P., Blond, S., David, R. ve Riogard, P. (2015). Anatomy, physiology and neurobiology of the nociception: A focus on low back pain (part A). Neurochirurgie, 22–S34, 22–34. doi:10.1016/j.neuchi.2014.09.001.
Mtui, E. G. (2016). Fitzgerald's Clinical Neuroanatomy and Neuroscience (7 b.). Elsevier.
Nagasako, E., Oaklander, A. ve Dworkin, R. (2003). Congenital insensitivity to pain: an update. Pain, 101, 213–9.
Nişanyan, S. (2021). Ağrı. Nişanyan Sözlük Çağdaş Türkçenin Etimolojisi. Erişim tarihi: 26.05.2021 https://www.nisanyansozluk.com/?k=a%C4%9Fr%C4%B1.
Park, T., Lu, Y., Jüttner, R., Smith, E., Hu, J., Brand, A. ve Lewin, G. (2008). Selective inflammatory pain insensitivity in the African naked mole-rat (Heterocephalus glaber). PLOS Biology, 6(1). doi:10.1371/journal.pbio.0060013.
Patterson, L., Dick, J. T. ve Elwood, R. W. (2007). Physiological stress responses in the edible crab. Cancer pagurus, to the fishery practice of declawing. Marine Biology, 152, 265-272.
Petersen-Felix, S. ve Arendt-Nielsen, L. (2002). From pain research to pain treatment: the role of human experimental pain models. . Best Practise and Research Clinical Anaesthesiology, 16(4), 667-680. doi:10.101053/bean.2002.0258.
Robblee, J. ve Starling, A. J. (2019). SEEDS for success: Lifestyle Management in Migraine. Cleveland Clinic Journal of Medicine, 86(11). doi:10.3949/ccjm.86a.19009.
Rose, J. D. (2002). The Neurobehavioral Nature of Fishes and the Question of Awareness and Pain. Reviews in Fisheries Science, 10(1), 1–38.
Smith, E. S. ve Gary, R. L. (2009). Nociceptors: a phylogenetic view. Comp Physiol A, 195, 1089–1106. doi:10.1007/s00359-009-0482-z.
Sneddon, L. U., Braithwaite, V. ve Gentle, M. (2003). Do fishes have nociceptors? Evidence for the evolution of a vertebrate sensory system. Proc. R. Soc. Lond. B, 1115-1121.
Sneddon, L. U., Elwood, R. W., Adamo, S. A. ve Leach, M. C. (2014). Defining and assessing animal pain. Animal Behaviour, 97, 201-212.
Sneddon, L. U. (2019). Evolution of nociception and pain: evidence from fish models. Phil. Trans. R. Soc. B 374: 20190290. http://dx.doi.org/10.1098/rstb.2019.0290.
Stephanie, Y. (2008). Fish and Pain Perception.https://www.wellbeingintlstudiesrepository.org/hsus_reps_impacts_on_animals/6 adresinden alındı.
Stevens, E. D. (2009). "Pain" and analgesia in fish: what we know, what we do not know, and what we need to know, before using analgesics in fish. Conference: Blue sky to deep water: the reality and the promise. ANZCCART Proceedings, (s. 115-124). Wellington, New Zealand. http://www.royalsociety.org.nz/organisation/panels/anzccart/archives/conferences/2008/ adresinden alındı.
The International Association for the Study of Pain (IASP). (2021). IASP Terminology. Erişim tarihi: 25.05.2021 https://www.iasp-pain.org/Education/Content.aspx?ItemNumber=1698.
The Senate Standing Committee on Legal and Constitutional Affairs. (2021). Do Invertebrates Feel Pain? . Erişim tarihi: 20.05.2021. https://sencanada.ca/content/sen/committee/372/lega/witn/shelly-e.htm.
Thompson, B. (2018). The Grasshopper Mouse and Bark Scorpion: Evolutionary Biology Meets Pain Modulation and Selective Receptor Inactivation. Journal of Undergraduate Neuroscience Education, Spring , R51-R53.
Walters, E. ve Williams, A. (2019). Evolution of mechanisms and behaviour important for pain. Phil. Trans. R. Soc. B, 374: 20190275. http://dx.doi.org/10.1098/rstb.2019.0275.
Wictionary The Free Dictionary (2021). Pain. Erişim tarihi: 26.05.2021. https://en.wiktionary.org/wiki/pain.
Yue, S. (2008). Fish and Pain Perception. Impacts on farm animals., 6. https://www.wellbeingintlstudiesrepository.org/hsus_reps_impacts_on_animals/6 adresinden alındı.